ГЕНЕТИКА ПОРОДЫ (ПОПУЛЯЦИИ)
Количество генов, определяющих признаки и свойства животного, поистине колоссально в любом живом организме. Тем не менее весь этот набор поддается определенной систематизации. Одна его часть определяет принадлежность особи к живой природе, и она, очевидно, одинакова или близка у всех живущих на земле. Другие части набора генов устанавливают принадлежность организма к тому или иному биологическому подразделению - типу, классу, семейству, роду и виду. В каждом биологическом подразделении у всех его представителей часть генов из общего набора близка между собой и свойственна представителям именно этого подразделения. Охотничьи собаки относятся, как и все другие, к виду Canis familiaris и несут набор генов, включающий подавляющую часть наследственных признаков, общих для всех представителей этого вида.
Однако все количество генов у собак не исчерпывается этим видовым набором. Существенное значение для племенной работы с определенной породой имеют те гены или их наборы, от которых зависят породные особенности. Что же они собой представляют? Мы знаем, что однопородные собаки обладают рядом породных признаков. Следовательно, имеется породный набор генов, свойственный всем особям одной породы. Кроме того, существуют различия между внутрипородными группами или типами. Это свидетельствует о наличии еще одного или нескольких «слоев» генов внутри породы. Наконец, индивидуальные различия между особями одной породы определяют соответствующие гены. Вся сумма генов, характеризующая породные, типичные и индивидуальные признаки одной породы и способная к образованию различных генных комбинаций, составляет ее генофонд и является основой племенной работы.
Генетика породы в отличие от генетики отдельной собаки изучает состояние и изменения всего генофонда. Поэтому на первый план выступают не простые качественные выражения тех или иных признаков единичной особи, а их количественное распределение между генами популяции. Большее значение приобретает и влияние внешних признаков факторов. Все это требует привлечения значительно более сложного математического аппарата, в частности методов статистического и вариационного анализа.
В современных условиях в связи с трудностями обмена кровями между отдельными удаленными друг от друга крупными кинологическими центрами большинство пород собак разводится и функционирует как набор более или менее изолированных групп (популяций), число особей в которых ограничено. Популяция - это группа однопородных животных, связанных между собой процессами размножения, т. е. непрерывного обмена генетическим материалом.
Собаки одной породы, разводимые в разных странах, могут заметно отличаться друг от друга. Поэтому при обсуждении задач конкретного разведения приходится говорить не о генетике породы в целом, а о генетике популяции и считаться с тем, что статистическая обработка данных из-за недостаточной численности не всегда точна.
Фактором, характеризующим генетическое состояние популяции, является частота генов, несущих признаки, в ее генофонде. В зависимости от частот отдельных генов в популяции складывается и соотношение генотипов и фенотипов. Под частотой генов понимают долю каждого аллеломорфного гена или аллеля, когда сумма всех имеющихся в популяции генов этого локуса при равнена к единице. Так, для пары аллелей Z и z, определяющих генотипически обусловленный признак и встречающихся в популяции с частотой р и q, где р = 1 - q, при помощи решетки Пеннета можно получить следующую генетическую формулу потомства первой генерации: P2ZZ + 2pqZz + q2zz.
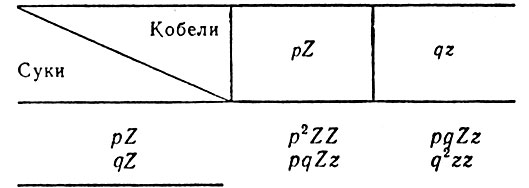
Генотипы первой генерации в популяции с р-частотой генов Z и q-частотой генов z
Это выражение называют законом Харди - Вейнберга. Оно представляет собой обычное разложение бинома (p+q)2 и устанавливает частоту наличия всех трех генотипов ZZ, Zz, zz в популяции, находящейся в стационарном, или равновесном, состоянии. Механизм, приводящий популяцию в такое состояние, заложен в свободном (случайном) скрещивании ее членов между собой и отсутствии отбора, одинаковой плодовитости членов популяции и некоторых других условиях. Стационарность, или равновесность, популяции в этом случае связана с тем, что частоты генов не меняются от поколения к поколению.
Степень отбора в охотничьем собаководстве такова, что по многим признакам (исключая дисквалифицирующие пороки) большинство пород или популяций очень близко отвечают этой модели. Это позволяет в первом приближении использовать закон Харди - Вейнберга для генетического анализа популяций. Наиболее просто устанавливается частота признаков, определяемых единичным геном. Зная число выщепившихся в популяции рецессивных гомозигот, например длинношерстных лаек (типа оленегонных) или рыжих гордонов, родившихся от нормальных родителей, можно выявить частоту появления этих особей и рассчитать число гетерозиготных по этому признаку собак в популяции, содержащих его в скрытой форме.
Частоты генотипов дигибридного признака, обусловленного двумя парами независимых генов А и а, В и в с частотами соответственно р и q, r и s, в стационарной популяции будут определяться произведением биномов (p + q)2 x (r + s)2.
Помимо характеристики по частотам генов единичных признаков, популяции свойственно соотношение генотипических и фенотипических групп, количественно выраженные признаки которых контролирует широкий набор генов. При таком наследовании частота и степень выраженности признака изображаются кривой с максимумом, лежащим между координатами родителей. Это соотношение, известное как закон Нильссона - Эле, для популяции должно быть дополнено следующими данными: отдельные гены, входящие в группу, контролирующую признак, могут отличаться степенью доминирования; возможно взаимодействие генов в контролирующей группе, изменяющее их влияние на признак; характер действия генов может в большей или меньшей степени зависеть от внешней среды.
Для характеристики популяции в случае полигенно-обусловленного признака ее разбивают на классы или группы по степени выраженности признака. Чем большее число генов влияет на изучаемый признак, тем меньше различие между отдельными классами, что приводит в конечном счете к возникновению непрерывной нормальной кривой (рис. 26). При доминировании число классов уменьшается.
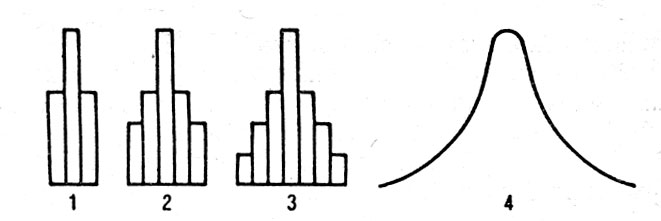
Рис. 26. Распределение частот генотипов Fsub2/sub при различных вариантах скрещивания: 1 - моногибридное; 2 - дигибридное; 3 - тригибридное; 4 - полигибридное
Генетическая характеристика популяции по количественным признакам вытекает из симметричного или асимметричного характера кривой. Определяют частоту и выраженность количественных признаков статистическими методами, которые дают возможность охарактеризовать популяцию по этим параметрам. Следует отметить, что ускорение отбора и консолидация популяции по одному какому-либо признаку, как правило, ведет к увеличению разброса по другим. Это объясняется тем, что число действующих генов, определяющих все свойственные породе признаки, крайне велико. Отбор на гомозиготность по одному из них не только не гарантирует повышения гомозиготности по другим, но зачастую дает обратную картину.
Исчезновение стационарности состояния в популяции происходит при воздействиях на нее. Частота генов может меняться от введения в популяцию собак других кровей. При этом изменение частоты Δр уже имеющегося в популяции гена р будет зависеть от исходной частоты гена р0, частоты этого гена у вводимых особей pi и доли популяции b, представленной вводимыми собаками. Изменение частоты гена рассчитывают так: Δр = b (pi - р0).
Влияет на частоту генов и так называемый дрейф генов. Его проявление связано с численностью популяции; в малочисленной популяции увеличивается фактор случайности в распределении отдельных генотипов. Ряд генов может вообще исчезнуть из генофонда в связи с отбраковкой их носителей. Другие гены могут накопиться в определенной части популяции. В случае увеличения численности случайно сложившиеся отношения могут резко расширить свою частоту и привести к быстрому возрастанию частоты отдельных генотипов.
Наиболее важным фактором, изменяющим структуру популяции, является отбор, который может быть направлен на исключение того или другого генотипа. Степень отбора, т. е. жесткость отбраковки, определяется коэффициентом отбора s. При s = 1 бракуемая группа исключается из разведения полностью; при s = 0,5 - на 50 % и т. д. Если исключению подвергается рецессивный генотип zz, то уменьшение частоты гена z будет составлять
| Δq= | sq2(1-q) | . |
| 1-sq2 |
Величина Δq зависит от величины коэффициента s, первоначального значения q и характера взаимоотношения аллелей Z и z, т. е. от полного или неполного доминирования. В собаководстве s = 1 имеет место только при выбраковке собак с дисквалифицирующим признаком (крипторхизм, неправильный прикус, непородность окраса или типа шерстного покрова и т. п.). Чаще величина s не бывает больше 0,4.
В соответствии с принятыми нормами экспертизы собаки с отклонениями от правильного полового типа (суки в «кобелиных ладах» и кобели в «сучьих ладах») получают оценку экстерьера не выше «хорошо». В результате к племенному использованию допускаются все суки, т. е. 50 % поголовья, и около половины кобелей, т. е. еще 25 %, так как кобели с такой оценкой используются не полностью. Таким образом, присуждение собакам с каким-либо нежелательным признаком оценки «хорошо» означает, что отбраковка этого признака производится с s = 0,2 / 0,3. При такой степени отбора снижение частоты гена q незначительно и составляет 0,001 - 0,01. Существенно зависит Δq от величины исходной частоты гена q. Расчет показывает, что изменение величины q низки как при больших, так и малых его значениях и наибольшие при средних (рис. 27).
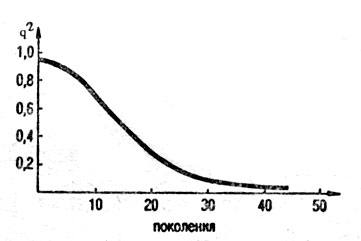
Рис. 27. Изменение частот рецессивного гена при отборе с коэффициентом 0,2 (по Д. С. Фолкнеру, 1985)
Скорость удаления из породы нежелательного признака продемонстрируем следующими примерами. При выбраковке с 5 = 0,2 снижение частоты гена от значения 0,8 - 0,9 (80 - 90%) до 0,1 (10%) может произойти за 50 - 60 поколений. Даже при полной выбраковке собак с дисквалифицирующим признаком (5=1) потребуется 10 поколений, чтобы снизить его частоту от 0,1 (10%) до 0,05 (5%), и еще 100 поколений - с 0,01 (1%) до 0,005 (0,5%). Отсюда становится понятным длительное сохранение в породах охотничьих собак рецессивных признаков, таких, как длинношерстность у западносибирских лаек, бронзовый оттенок у черно- и кофейнопегих пойнтеров, короткошерстность у дратхааров и т. п.
В приведенных примерах речь шла об отборе, направленном на ликвидацию рецессивного признака. При отборе против доминантного признака носители его обнаруживаются сразу, и гомо- и гетерозиготные по соответствующему гену особи могут быть отбракованы с s = 1. При этом уже в первом поколении F1 частота рецессивного гена достигнет единицы. Таким образом, в животноводстве действует парадоксальное только на первый взгляд положение, что вести породу гораздо легче по рецессивному признаку. Вспомним, что, по имеющимся данным, первоначальные средневековые легавые имели бурый или кофейный рецессивный окрас. Очевидно, легкость отбора по этому признаку и невозможность его изменения без прилива чужой крови способствовали формированию средневековых легавых.
Необходимо остановиться на некоторых особенностях малочисленных популяций, которые широко представлены в породах охотничьих собак. Как указывалось выше, в таких популяциях существенное значение приобретает дрейф генов, который может привести к утере в полуляции части генофонда. В силу случайности таких утрат они в разных популяциях одной и той же породы могут касаться различных составных частей генофонда. Это приводит к возрастанию обособленности популяций и увеличению различий между входящими в них собаками. В многочисленных популяциях этот процесс проявляется слабее.
Другая особенность малочисленных популяций - неизбежность возрастания в них инбридинга, без которого такая популяция не может существовать. Это ведет к увеличению в них числа гомозиготных по отдельным признакам собак и одновременно к расщеплению популяции.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://kinlib.ru/ 'Библиотека по собаководству'